Plastizität von Herzmuskelzellen des Molches
Forschungsbericht (importiert) 2006 - Max-Planck-Institut für Herz- und Lungenforschung - W. G. Kerckhoff-Institut
Nach wie vor sind Erkrankungen des Herz-Kreislaufsystems in den Industrieländern mit Abstand die häufigste Ursache für einen vorzeitigen Tod. So vielfältig die Erkrankungen sein können, führen sie jedoch in den meisten Fällen zu einem Verlust funktionellen Herzgewebes und einer Vernarbung ehemaliger Herzmuskulatur. Obwohl Säugetiere wie der Mensch durchaus in der Lage sind, gewisse Organe wie Leber und Skelettmuskeln nach einer Schädigung wiederherzustellen, ist die Fähigkeit, den Herzmuskel nach einer Schädigung zu regenerieren, nicht oder nur in sehr geringem Maße vorhanden.
Es ist deshalb von vorrangigem wissenschaftlichen Interesse, Therapien zu entwickeln, die zu einer nicht nur messbaren, sondern für den Patienten auch spürbaren Verbesserung führen. Vielfältige therapeutische Ansätze befinden sich bereits im Stadium klinischer Erprobung; sie stützen sich in vielen Fällen auf die Gabe von Stammzellen unterschiedlicher Herkunft oder auf die Stimulation proliferativer Vorgänge im Herzmuskel [1; 2; 3; 4]. Obwohl einige dieser Studien durchaus messbare Verbesserungen der Herzfunktion aufzeigen, konnten sie die gesteckten Ziele bisher nicht erreichen.
Die Fähigkeit zur Regeneration ist unter den eukaryontischen Organismen weit verbreitet. Während primitive Organismen – wie Nesseltiere und Flachwürmer – aus Teilen des Körpers einen ganzen Organismus rekonstituieren können, ist die Fähigkeit der Regeneration multipler Organe in Wirbeltieren nur bei wenigen Organismen beschrieben [5] . Die Meister der Regeneration unter den Wirbeltieren sind zweifelsohne unter den Amphibien zu finden [6]. Der Modellorganismus, der am MPI für Herz- und Lungenforschung in Bad Nauheim verwendet wird, ist der grünliche Wassermolch Notophthalmus viridescens. Dieser Molch besitzt nicht nur die Fähigkeit, komplette Gliedmaßen nach Amputation zu regenerieren, sondern auch die nahezu einzigartige Eigenschaft, Teile des Zentralnervensystems sowie das Herz nach Amputation oder mechanischer Schädigung wiederherzustellen. Ein fundamentaler Unterschied besteht in der Teilungsfähigkeit der Herzmuskelzellen des Molches, welche Säugetieren nicht zu eigen ist. Am MPI in Bad Nauheim wollen die Wissenschaftler verstehen, welche Plastizität Herzmuskelzellen aus dem Molch besitzen, sprich, wie wandlungsfähig diese Zellen sein können und welche molekularen Mechanismen die Umkehr zelltypspezifischer Eigenschaften (Dedifferenzierung) und Teilungsfähigkeit unterstützen.

Teilungsfähigkeit von Herzmuskelzellen des Molches
Jeremy Brockes und Kollegen konnten zeigen, dass kultivierte Herzmuskelzellen (Kardiomyozyten) aus ungeschädigten Molchherzen eine gewisse Heterogenität in ihrer Proliferationsfähigkeit besitzen [7]. Es wurde beobachtet, dass manche Kardiomyozyten mehrere Teilungszyklen durchlaufen können, während andere sich in Kultur nicht teilten. Die Frage, ob diese Heterogenität auf unterschiedliche Zellpopulationen zurückzuführen ist oder ob sich die Kardiomyozyten durch die Anzahl der durchlaufenen Zellzyklen unterscheiden, konnte diese Studie nicht klären.
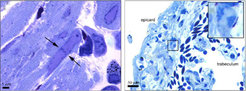
Primäres Ziel der Forscher am MPI für Herz- und Lungenforschung war zu untersuchen, wie sich Kardiomyozyten in vivo verhalten [8]. In ungeschädigtem Herzen konnten die Wissenschaftler zwar eine substantielle Anzahl an Kardiomyozyten beobachten, die eine Kernteilung zeigen, aber es fanden sich nur sehr wenige Kardiomyozyten, die auch eine Zellteilung durchführten. Nach einer Schädigung des Herzens wurden jedoch sehr viele sich teilende Zellen beobachtet, die teilweise auf Kardiomyozyten zurückzuführen waren, was Reste sarkomerer Strukturen bewiesen (Abb. 1). Zusätzlich konnten die Forscher einen rapiden Abbau sarkomerer Strukturen in kurz zuvor geschädigten Herzen beobachten, was mit einer massiven Reduktion der Expression von herzmuskelspezifischen Proteinen, wie beispielsweise der schweren Kette von alpha-Myosin oder herzspezifischem Troponin T, einherging (Abb. 2). Eine Reexpression dieser Marker erfolgte erst zu späteren Zeitpunkten der Regeneration nach etwa 14 Tagen, was darauf hindeutete, dass für regenerative Vorgänge ein transienter Umbau zellulärer Strukturen stattfinden muss.

Verhalten von Kardiomyozyten in Kultur und ungeschädigter Umgebung
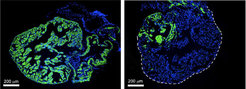
Isolierte Kardiomyozyten exprimieren herzspezifisches Troponin T und die schwere Kette von alpha-Myosin über die Dauer der Kultur hinweg, zusätzlich beginnen die Herzmuskelzellen nach 6 bis 8 Tagen zu schlagen, was einen Hinweis darauf gab, dass der differenzierte Phänotyp dieser Zellen unter Kulturbedingungen erhalten blieb.
Nach Markierung der Kardiomyozyten mit dem membranständigen Farbstoff Dil wurden diese in intakte Hinterbeine von Molchen injiziert. 8 und 15 Tage nach Injektion behielten die transplantierten Kardiomyozyten ihr charakteristisches Aussehen und exprimierten nach wie vor kardiale Marker, was darauf hindeutete, dass sie auch in dieser fremden Umgebung ihre differenzierte Identität behalten.
Umprogrammierung von Kardiomyozyten im regenerativen Umfeld
Um das Verhalten von isolierten Kardiomyozyten in einer regenerierenden Umgebung zu untersuchen, wurden diese in einen regenierenden Zellverband im Beinstumpf, ein sogenanntes Blastem, 5 Tage nach Amputation injiziert. In dieser Umgebung findet – ähnlich wie beim geschädigten Herzen – eine rapide Abnahme sarkomerer Strukturen in den Kardiomyozyten statt, welche auch 7 Tage nach Transplantation bestehen bleibt. Obwohl die markierten Zellen im geschädigten Gewebe verbleiben, findet eine Reexpression der kardialen Marker auch 15 bzw. 30 Tage nach Injektion nicht statt. Lediglich die Expression des Intermediärfilament-Proteins Desmin, welches in gestreiften und glatten Muskelzellen vorkommt, blieb erhalten. Wurden die Kardiomyozyten in geschädigte Herzen injiziert, so verloren sie ebenfalls ihren differenzierten Phänotyp, was darauf hindeutet, dass sowohl orthotopische wie auch heterotopische regenerative Umgebungen die Dedifferenzierung von Kardiomyozyten einleiten können.
Dedifferenzierung von transplantierten Kardiomyozyten
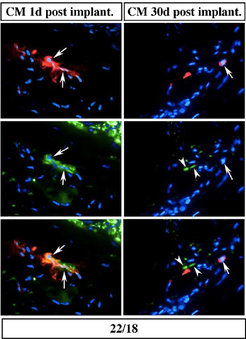
Um herauszufinden, ob das Abschalten kardialer Marker mit dem Anschalten von Markern für undifferenzierte Zellen einhergeht, wurden regenerierende Beinstümpfe gegen den Blastemmarker 22/18 gefärbt (Abb. 3). Tatsächlich exprimieren etwa ein Drittel der transplantierten Kardiomyozyten 1 Tag nach Transplantation den Blastemzellmarker, welcher 30 Tage nach Transplantation nicht mehr in den Kardiomyozyten nachweisbar ist.
. Nach 30 Tagen sind diese Proteine nicht mehr nachzuweisen (rechts). Transplantierte Herzmuskelzellen sind rot gefärbt, das Protein 22/18 grün und die Zellkerne blau.")
Molch-Kardiomyozyten verwandeln sich in regenerierendem Beinstumpf hauptsächlich zu Skelettmuskelzellen
Umgekehrt zur Abnahme der Expression von Blastemmarkern in transplantierten Kardiomyozyten findet eine Expression von skelettmuskelspezifischen Proteinen, wie beispielsweise sarkomerisches Myosin schwere Kette und Skelettmuskel Troponin I, ab etwa 15 Tagen nach der Transplantation statt. Nach etwa 30 Tagen formierten sich die ehemaligen Kardiomyozyten zu kleinen Muskelfasern, die auch wiederum positiv für Skelettmuskel-Troponin I waren (Abb. 4). Dies ist bei etwa 60% aller DiI-positiven Kardiomyozyten der Fall. In den transplantierten Herzmukelzellen finden sich zu diesem Zeitpunkt keine Spuren mehr von herzspezifischen Proteinen.
. Bereits 2 Tage nach Transplantation werden herzmuskel-spezifische Proteine in den Kardiomyozyten abgeschaltet. Die transplantierten Kardiomyozyten sind rot markiert, die Immunfärbung gegen skelettmuskelspezifisches Troponin I (links) und herzmuskelspezifisches Troponin T (rechts) sind grün markiert. Die Zellkerne wurden blau gefärbt.")

Die Teilungsfähigkeit der Kardiomyozyten setzt keine Dedifferenzierung voraus
Die Max-Planck-Wissenschaftler färbten dazu kultivierte Molchkardiomyozyten gegen phospho-Histon H3, einen Marker für die G2-Phase des Zellzyklus. Im Verlaufe der Kultivierung zeigten immer mehr Herzmuskelzellen posphoryliertes Histon H3, jedoch exprimierten diese Zellen gleichzeitig immer noch Myosin schwere Kette, was darauf hinweist, dass die Fähigkeit zur Proliferation nicht notwendigerweise mit einer Dedifferenzierung der Zellen verknüpft ist. (Abb. 5). Eine Färbung gegen phosphoryliertes Histon H3 in regenerierendem Beinstumpf detektierte ebenfalls positive Signale in transplantierten Kardiomyozyten, wodurch bestätigt wurde, dass ehemalige Kardiomyozyten zur Zellteilung fähig sind, unabhängig davon, ob sie eine Identität als reife Herzmuskelzelle oder einen dedifferenzierten Status annehmen.
Borchardt und sein Team konnten mit dieser Studie zeigen, dass Molchkardiomyozyten, wenn man sie in ein regeneratives Umfeld transplantiert, eine bemerkenswerte Plastizität aufweisen. Das drückt sich nicht nur durch die Expression typischer Markerproteine des Zielgewebes aus, sondern auch in der Beteiligung am Wiederaufbau der Zielstruktur, in diesem Falle der Skelettmuskulatur. Kardiomyozyten aus dem Molch besitzen zudem die Eigenschaft, sich in blastemartige Zelltypen zu verwandeln, wenn sie Signale aus einem regenerierenden Gewebe erhalten. Die Tatsache, dass kultivierte Kardiomyozyten in den Zellzyklus eintreten können, steht im Gegensatz zu regenerierenden Herzen und zu Kardiomyozyten, die in geschädigte Beinstümpfe implantiert wurden, und zeigt hiermit, dass der Eintritt in den Zellzyklus und eine Dedifferenzierung nicht notwendigerweise miteinander einhergehen.
 als auch in regenerierendem Gewebe teilen (rechts). Kardiomyozyten wurden rot markiert, die Immunfärbung gegen phoshoryliertes Histon-H3, einen Indikator für Zellen, die sich in der G2-Phase des Zellzyklus befinden, ist grün markiert, die Zellkerne wurden blau gefärbt.")

Zukunftsaussichten
Die Wissenschaftler am MPI für Herz- und Lungenforschung sind auf der Suche nach den exakten Signalen, die eine Gewebeschädigung mit einer Regeneration verbinden und damit verhindern, dass ein geschädigtes Gewebe vernarbt oder gar abstirbt. Hierzu haben sie Projekte initiiert, welche die Expression von Genen zu unterschiedlichen Zeitpunkten der Regeneration mit der im ungeschädigten Herzen vergleicht. Es ist weiterhin nicht geklärt, ob Kardiomyozyten allein oder im Zusammenspiel mit anderen Zelltypen den Wiederaufbau des Herzmuskels bewerkstelligen. Zudem etablieren die Forscher primäre Zellkulturen aus regenerierenden Herzen, um zu beantworten, welche Identität Zellen des ungeschädigten und geschädigten Herzens besitzen und welche Verwandlungsfähigkeit diesen verschiedenen Zelltypen eigen ist. Wenn man die molekularen Vorgänge der Herzregeneration im Molchherzen genauer charakterisiert hat, kann dies einen großen Schritt in die Richtung Selbstheilung von Säugetierherzen bedeuten.